Mechanisms of memory, selection and consolidation
2026年5月25日上午,北京脑科学与类脑研究所邀请纽约大学格罗斯曼医学院(NYU Grossman School of Medicine)György Buzsáki 教授开展了一场题为 “Mechanisms of memory, selection and consolidation” 的学术报告。报告由本所研究员仵婷主持。

Dr. György Buzsáki
Biggs Professor of Neuroscience, Department of Neuroscience at NYU
Dr. György Buzsáki于1974年在匈牙利佩奇大学(University of Pécs, Hungary)获得医学博士,1984 年在布达佩斯匈牙利科学院(Academy of Sciences, Budapest)获得神经科学博士。其在匈牙利佩奇大学攻读博士阶段师从匈牙利著名神经生理学家Endre Grastyán(1929–1992),Grastyán关注于海马生理、脑节律和行为-认知关系的研究,对Buzsáki后续关于海马 theta、gamma、sharp-wave ripples、神经序列和记忆巩固的研究产生了重要影响。Buzsáki还曾于1980-1981年在University of Texas, San Antonio进行神经科学博士后训练,1981-1982年在University of Western Ontario, London进行博士后训练。
Dr. György Buzsáki的研究围绕“神经句法”(neural syntax)这一概念展开,即认为脑功能不仅受外界刺激反应所支配,也受到由脑节律产生的、内部生成的、类似规则的时间结构所调控。他的研究重点之一是海马-皮层网络。在这一领域,他推动阐明了 theta节律、gamma节律以及睡眠相关的尖波—高频振荡(sharp wave–ripples)等神经振荡,如何协调细胞集群和序列活动,并进一步支撑学习、记忆和导航等认知过程。他尤其以“两阶段记忆巩固模型”(two-stage model of memory consolidation)而著称,该模型强调学习后的睡眠期再激活,尤其是与尖波—高频振荡事件相关的再激活,对于长期记忆的稳定化具有关键作用。Buzsáki 的研究为理解神经振荡、同步化活动以及跨频耦合如何塑造健康大脑和神经系统疾病中的信息处理过程,提供了一个统一性的理论框架。
报告开头,Dr. Buzsáki以动物在自然环境中的觅食和导航情境作为引入:动物观察到远处线索后,需要推断目标位置,同时绕开河流、树木或其他障碍物。这个例子揭示出动物的行为并不是简单的刺激—反应,而是包含场景维持、目标预测、路径规划和行动后果评估的复杂过程。Dr. Buzsáki随后提出,大脑演化的核心目标之一并非被动再现外部世界,而是预测行动后果(predict the consequences of actions)。即使是简单生命体,也需要维持自身内部动态、产生动作输出,并记录动作带来的结果。随着神经系统复杂化,大脑可以在更长时间尺度和更复杂环境中进行预测,例如决定是否前往某个地点、如何规避风险、如何根据过去经验计划未来行动。因此,记忆并不是静态存储,而是服务于行动预测和未来规划的动态结构。
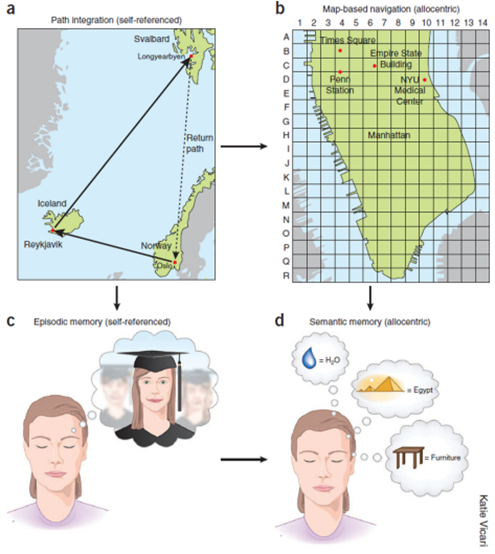
Buzsáki & Moser, Nature Neuroscience, 2013
报告随后讨论海马功能的经典争议。心理学传统通常将hippocampus视为记忆系统的核心结构,尤其与情景记忆(episodic memory)相关;而神经生理学传统则强调海马中的place cells,将海马视为大脑的空间导航系统或“GPS”。Dr. Buzsáki提出,真正的问题并不是二者谁对谁错,而是:空间导航和记忆是否共享同一类神经计算机制?
Dr. Buzsáki将导航分为两类。第一类是self-referenced / egocentric navigation,即以自身为参照的导航,依赖displacement、duration、velocity和path integration。动物即使在黑暗中,也可以通过自身运动信息估计返回原处。第二类是map-based / allocentric navigation,即基于外界坐标或地图的导航。在这种表征中,具体路径和时间细节被剥离,保留的是位置之间的关系。Dr. Buzsáki将这种从自我参照到客观地图的转化类比为从情景记忆到语义记忆的转化。在记忆层面,episodic memory是自我参照的,semantic memory则逐渐剥离具体时空背景,形成抽象知识。在上述基础上,Dr. Buzsáki提出“cognition is internalized action”的观点。大脑最初服务于身体在环境中的行动,但随着内部模型的发展,行动序列可以在没有实际动作输出的情况下被内部模拟。此时,记忆、想象和计划不再只是对过去的被动记录,而是对可能行动路径的离线运行。
Dr. Buzsáki详细介绍了其近期发表的Nature研究“Hippocampal neuronal activity is aligned with action plans”。该研究设计了一个acoustic cue-guided navigation task(以声频为线索的导航任务),以区分海马活动到底是由空间位置、声音频率、奖励线索驱动,还是由动物内部形成的行动计划驱动。
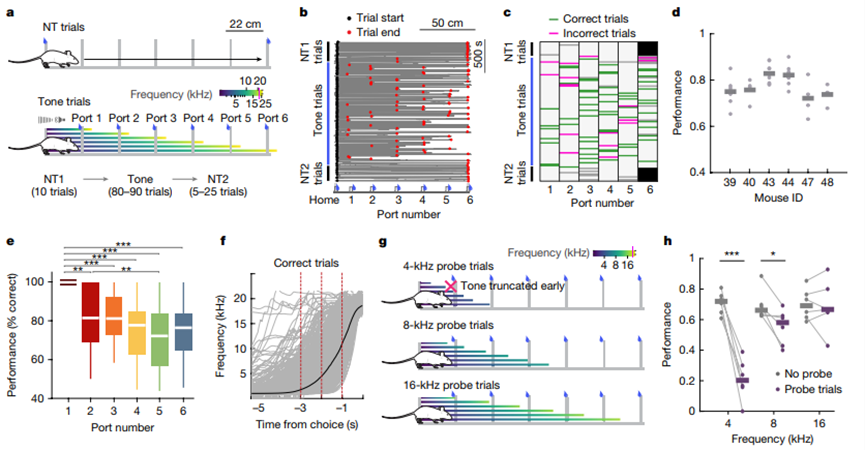
Zutshi et al.Nature, 2025
该任务中,动物从Home出发,在一条线性轨道上面对Port 1至Port 6等多个取水口。实验包括两类trial:首先是no-tone trials,动物在没有声音线索的情况下沿轨道往返;随后是tone trials,声音频率从约1 kHz 增加至22 kHz,用作目标位置或行动进程的线索。动物需要根据声频变化判断应该前往哪个port获得水奖励。实验的关键设计在于,研究者可以改变或截断声音线索,使空间、声音、奖励和行动计划之间不再完全重合,从而判断海马神经元究竟跟随哪个变量。
结果显示,海马神经元表面上可以表现出对空间位置、声音频率、奖励和动作状态的调谐。但当这些变量被实验性分离后,海马群体活动更接近于动物当前的online action plan(在线行动计划)。PPT《Spike sequences change perpetually even when environmental inputs remain constant》中展示,即使环境输入保持恒定,神经放电序列仍持续变化。这说明海马并不是外部世界的简单投影,而是在内部动力学、任务结构和目标计划的共同作用下生成序列。主讲人强调,当大脑“决定”一个新的计划时,海马群体活动轨迹也会随之改变。
这一发现挑战了将海马简单视作“空间GPS”的传统观点。更准确地说,海马不是只记录“动物在哪里”,而是同时组织“动物正在计划去哪里、接下来要做什么、当前经验处于任务进程的哪个阶段”。因此,place cells、time cells、tone-tuned cells或reward cells可能并非彼此独立的细胞类别,而是在不同实验结构下被行动进程“投影”出来的表观调谐。Dr. Buzsáki概括了神经元集群序列的两种产生方式。第一种是outside-in:外界线索、感觉输入、路径整合等上游机制依次驱动神经元群体放电。例如动物在真实轨道上移动时,外部线索和身体运动可以诱发一系列顺序激活的place cell assemblies。第二种是inside-out:神经系统依靠自身内部动力学、突触连接和延迟结构生成序列,即使没有直接外界输入,也能在静息、睡眠或规划状态下重新运行相似轨迹。实际行为中的神经序列可由外界输入诱发,但记忆重放、未来规划和想象则更多依赖inside-out 的内部动力学。Dr. Buzsáki强调,过去经验、未来想象和当前计划并不是三种完全分离的神经过程,而是同一种序列机制在不同时间方向上的表现。过去经验被重新激活时称为 memory replay;尚未执行的路径被内部运行时则可表现为prediction或planning。
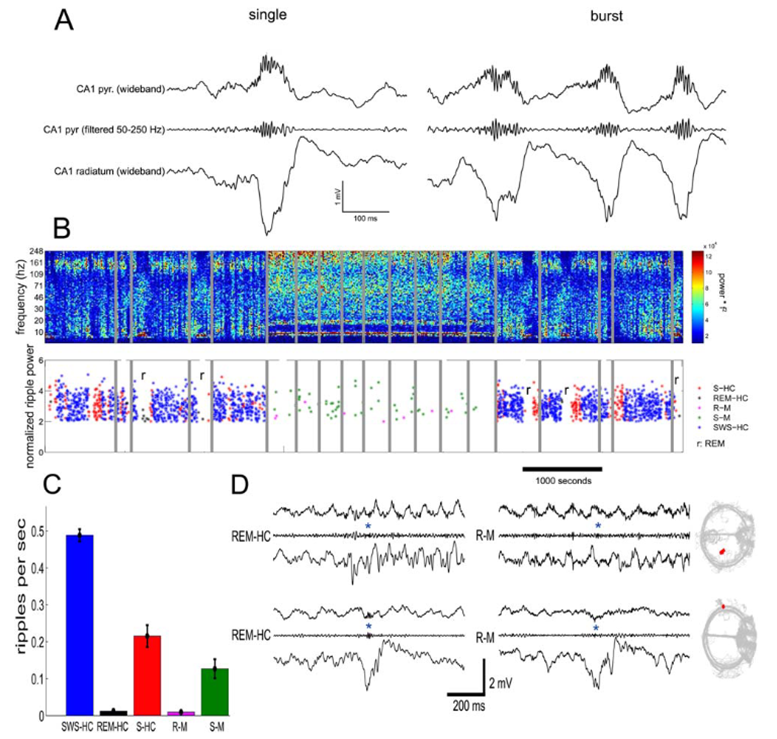
Buzsáki et al. Hippocampus, 2015
Dr. Buzsáki详细讲述了sharp-wave ripples(SPW-Rs)的机制部分。SPW-Rs 是海马中高度同步的群体活动事件,通常包括来自CA3到CA1的sharp wave,以及由CA1锥体细胞和中间神经元相互作用产生的ripple高频振荡。Dr. Buzsáki指出,SPW-Rs是正常脑中最同步、最强烈的群体活动模式之一,常出现在清醒静息、停顿状态和睡眠中,而不是动物主动探索外界的高唤醒状态。
在Buzsáki的理论框架中,SPW-Rs具有三方面关键功能。第一,它们能够压缩重放动物之前经历过的神经序列,使数秒甚至更长时间尺度的行为轨迹在约百毫秒内被快速重现。第二,它们可以在清醒静息期标记重要事件,为之后睡眠中的选择性巩固提供“权重”。第三,它们能通过海马-新皮层耦合影响更广泛脑区,推动系统层面的记忆巩固。
Dr. Buzsáki 的研究“Cholinergic suppression of hippocampal sharp-wave ripples impairs working memory”和“Interaction of acetylcholine and oxytocin neuromodulation in the hippocampus”展示了乙酰胆碱(ACh)、多巴胺(DA)和催产素(OXT)等神经调质系统与SPW-Rs的关系。Dr. Buzsáki强调,SPW-Rs并不是在所有脑状态下都会发生;它们更倾向于出现在低唤醒、静息和离线状态。当动物处于高度警觉、主动探索或强烈感觉输入状态时,胆碱能等神经调质系统会抑制SPW-Rs,从而让海马进入更适合在线编码外界输入的模式。这一观点说明,记忆编码与记忆巩固需要不同脑状态。清醒探索时,大脑更适合接收外界信息、形成新的行动计划;静息和睡眠时,大脑则减少外界驱动,使海马能够通过 SPW-Rs进行重放和整合。
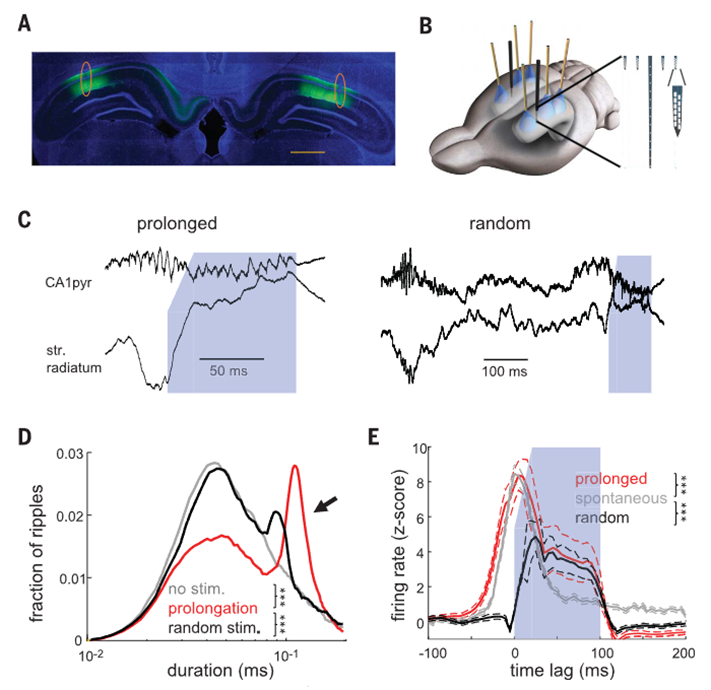
Fernández-Ruiz et al., Science, 2019
随后,Dr. Buzsáki进一步介绍了闭环光遗传实验。研究者实时记录海马CA1局部场电位,在线检测自然发生的SPW-R;一旦系统检测到SPW-R,就立即触发光遗传刺激,激活海马锥体神经元,从而将这次自然发生的ripple延长几十到约一百毫秒。关键在于,这不是随机照光或随机制造同步活动,而是顺着动物大脑自然启动的SPW-R进行闭环延长。对照条件包括no stimulation和random stimulation。行为结果显示,闭环延长自然发生的SPW-Rs可以提高动物在空间记忆任务中的表现,表现为学习曲线更快上升、最终正确率更高;而随机刺激不能产生相同效果。相反,如果实验性干扰SPW-Rs,动物第二天的记忆表现会下降。
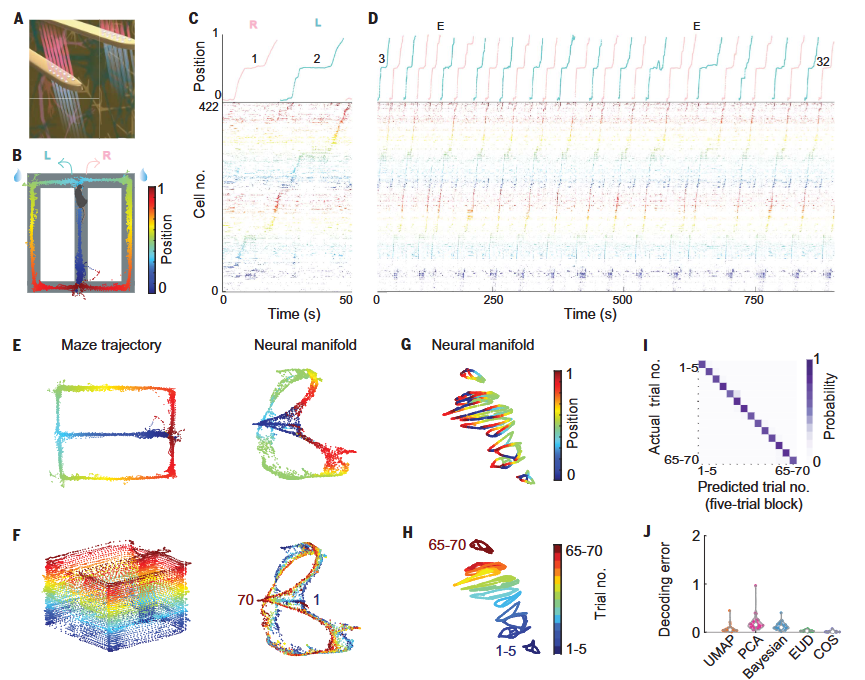
Yang et al., Science, 2024
在解释了SPW-Rs的因果作用后,Dr. Buzsáki转向更核心的问题:如果生活中有大量经验,为什么只有一部分进入长期记忆?这对应记忆选择(memory selection)问题。不同经历并不会被大脑同等标记;某些trial周围出现更多或更强的waking SPW-Rs,提示这些事件被赋予了更高的“重要性权重”。
报告中还讨论了海马与新皮层在记忆巩固中的分工。海马被描述为更可塑、更适合快速编码事件索引的区域;新皮层则更稳定,更适合长期保存抽象化和泛化后的表征。这对应经典的systems consolidation框架,但Dr. Buzsáki更强调其中的动态节律耦合:海马 SPW-Rs、新皮层慢振荡(slow oscillations)和丘脑-皮层纺锤波(sleep spindles)之间可能构成多层级的时间窗口,使海马重放能够在合适的时相影响皮层突触可塑性。报告最后讨论了稳定性-可塑性困境(stability-plasticity dilemma)。高稳定、低可塑会导致no learning;高可塑、低稳定会导致 interference;只有在二者之间达到balance,系统才能既学习新信息,又不破坏已有记忆。Dr. Buzsáki将这一问题与人工智能中的catastrophic interference / catastrophic forgetting类比,指出无论是生物脑还是人工神经网络,只要持续学习新任务,都必须解决新信息覆盖旧信息的问题。对于这一问题Dr. Buzsáki展示了未发表的发现,提出的解决方案是nested computation / division of labor。第一,脑区之间存在分工:海马更可塑,适合快速编码;新皮层更稳定,适合长期保存。第二,时间状态存在分工:清醒状态主要进行encoding,睡眠状态主要进行stabilization。第三,神经元群体也存在分工:高放电神经元可能更像 generalizers,负责抽象和泛化;低放电神经元可能更像specifiers,负责区分细节。第四,神经活动在population manifold 或computational subspaces中被组织,不同任务或记忆可以占据不同低维子空间,从而减少干扰。
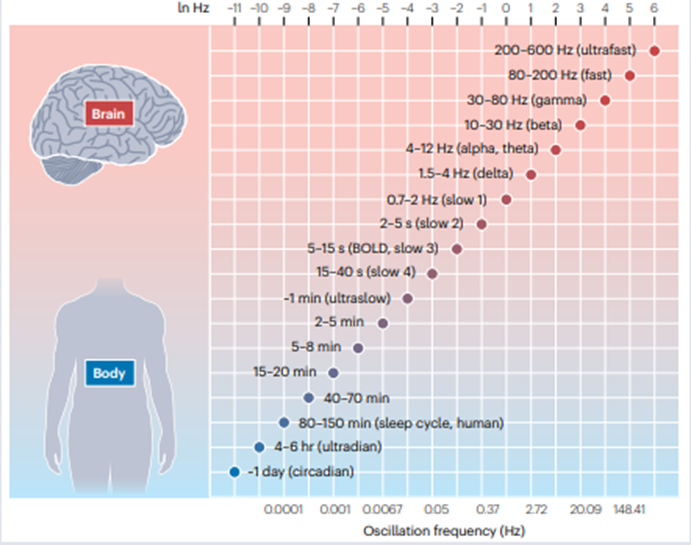
Buzsáki, Nat Rev Neurosci, 2026
Dr. Buzsáki 强调,大脑和身体并不是由单一频率组织,而是由一系列嵌套节律共同调控。这些节律之间的核心机制是phase-amplitude coupling(相位-振幅耦合):较慢节律的相位可以调制较快节律的幅度,从而为神经元放电、信息打包、跨区域通信和行为状态转换提供时间窗口。SPW-Rs 作为超快、高同步事件,必须嵌入到更慢的睡眠节律、皮层慢波和全身节律中,才能有效参与记忆选择和巩固。
报告的最后,听众对报告内容展开了深入友好的讨论。提出的问题包括行为学实验设计和对脑机接口的观点看法,Dr. Buzsáki 对各个问题一一作答。
撰稿人:杨子林
